Introduction
- Cytokines are a group of proteins and peptides that constitute the intercellular communication network of every cell system of the body, including the immune system.
- They function as signaling molecules to regulate the growth, differentiation, activation, and inhibition of all cellular aspects of both the innate and adaptive immune responses.
- Chemokines are a specialized subset of cytokines that function to induce directed cell movement, i.e., chemotaxis, in nearby responsive cells; they are chemotactic cytokines, hence the name “chemokines”.
- Chemokines recruit leukocytes to sites of inflammation and play an important role in lymphocyte trafficking.
- Cytokines and chemokines play a central role in cell development and differentiation, as well as inflammatory and immune responses from the initial induction of the innate immune response, progressing to the vast array of feedback signals and the DNA recombinatorial and mutational events involved in the adaptive immune system.
- Cytokines and chemokines are involved in maintaining balanced homeostasis of the innate and adaptive immune system in health and play critical roles in the pathogenesis of infectious diseases, allergic and autoimmune disorders, and in cancer.
- Cytokines are signaling molecules similar to hormones and neurotransmitters that enable one cell to communicate with another. Cytokines differ, however, from conventional hormones in several important ways (TABLE 1).
- For example, unlike hormones, which are produced as single cell products of the endocrine system, are present in plasma, and affect specific target cells, cytokines show pleiomorphism, i.e., they are produced by a variety of cells, are more localized to tissues, and affect a larger number of cells of the immune system.
- Secondly, cytokines show a high degree of redundancy in their biologic function, and different cytokines may have similar effects in contrast to hormones whose effects are more restricted.
- Cytokines also display a relatively higher degree of synergism than hormones so that their combined effects are greater than the sum of their individual effects.
[table id=150 /]
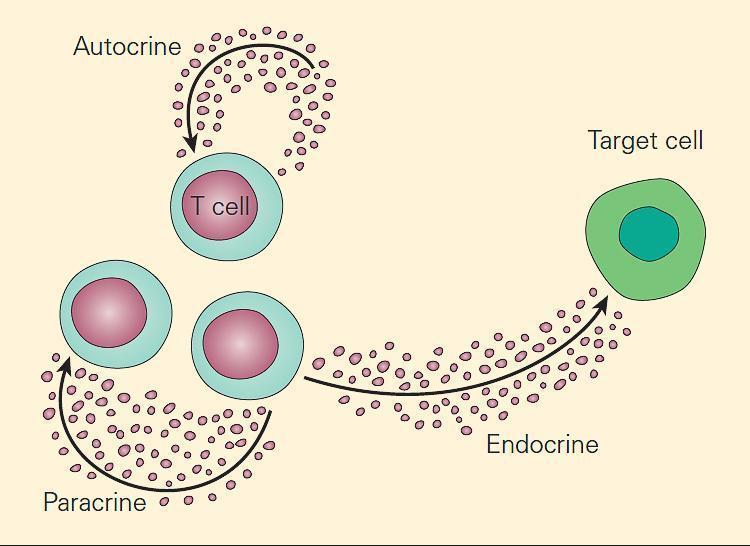
Despite their differences, cytokines are similar to hormones in their modes of action, including the following:
- autocrine, if the cytokine acts on the cell that secretes it,
- paracrine, if the action is restricted to the immediate vicinity of a cytokine’s secretion, and
- endocrine, if the cytokine diffuses to distant regions of the body (carried by blood or plasma) to affect different tissues (TABLE 1 and FIGURE 1)
Cytokine Classification
- Cytokines have been traditionally named according to their cell of origin or their specific functions.
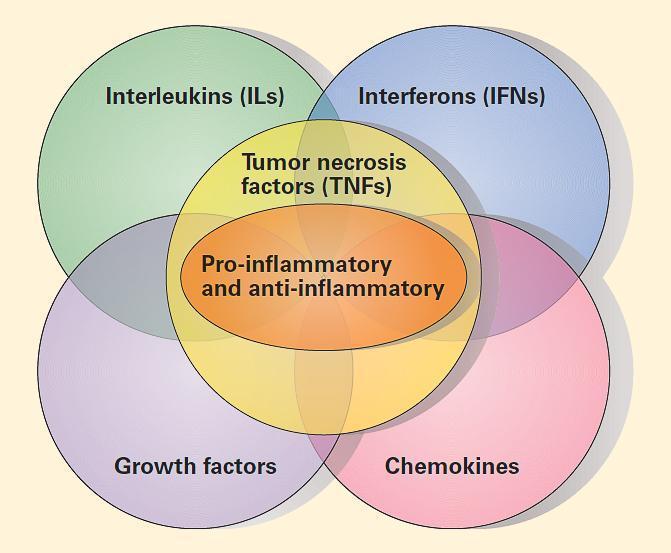
- TABLE 2 summarizes the major groups of cytokines: (1)interleukins (ILs); (2) interferons (IFNs); (3) pro-inflammatory and anti-inflammatory cytokines; (4) growth factors (GFs); and (5) chemokines.
[table id=151 /]
- TABLE 3 shows the functional classification of cytokines.
[table id=152 /]
- These proteins are a heterogeneous collection of cell-derived peptides with low molecular weights ranging from eight to 45 kDa that function as cell-to-cell messengers and exert their effects by binding to specific high-affinity receptors on cell surfaces.
- The elucidation of the molecular structure of cytokines and their receptors and signalling pathways has explained to some degree their pleiotropic properties and their redundancy.
- Cytokines and cytokine receptors consist of various protein subunits that are shared respectively with one another. The pleiotropic properties of a cytokine are related to its ability to bind to receptors that are found on multiple cellular lineages or by its ability to activate multiple signaling pathways that differentially contribute to different functions.
- Overlapping actions or redundancy are related to the finding that different cytokines can bind to the same or to a variety of receptors that share the same or similar molecular motifs.
The Balance of Pro-inflammatory Versus Anti-inflammatory Cytokines
- Cytokines can exert opposing (antagonistic) functions and are very efficient at maintaining the immunologic balance between the external and internal environments. Cytokines accomplish these antagonistic effects by either promoting or inhibiting inflammation (FIGURE 2).
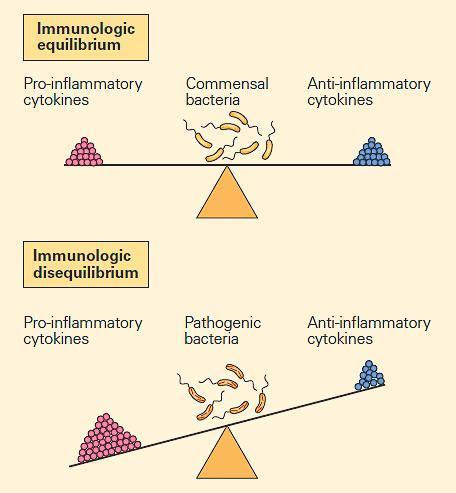
- This is accomplished because the same molecules can react with receptors that share homologies and that can be activated or turned off depending on the nature of the foreign insult and the resulting inflammatory response.
- FIGURE 3 shows a schematic representation of the overlapping nature of the major groups of cytokines. The resulting proinflammatory/anti-inflammatory paradigm is superimposed on this overlapping and cascading set of responses and is responsible for the expression of cytokine production in health and disease.
Effects of Cytokines on CD4+ T Cell Differentiation
- FIGURE 4, below, shows the vast repertoire of cytokines involved in CD4+ T cell differentiation.
- Following contact of APCs with naïve CD4+ T cells, the cytokine milieu that is produced is an important factor that initiates the process by which naïve CD4+ T cells begin to differentiate toward one of several subset fates.
- Signals from the TCR, as well as from IL-12 and IFN-γ (acting through STAT4 and STAT1, respectively), increase the expression of the transcription factor T-bet, which promotes IFN-γ production and commitment to the Th1 cell lineage.
- Naïve CD4+ T cells are induced to become Th2 cells through the secretion of IL-4 by innate immune cells and mast cells, which signal through STAT6.
- This leads to expression of the transcription factor GATA3, in turn resulting in the production of IL-4, IL-5, and IL-13, which are important for host defense against helminths and also contribute to the pathogenesis of asthma and allergy.
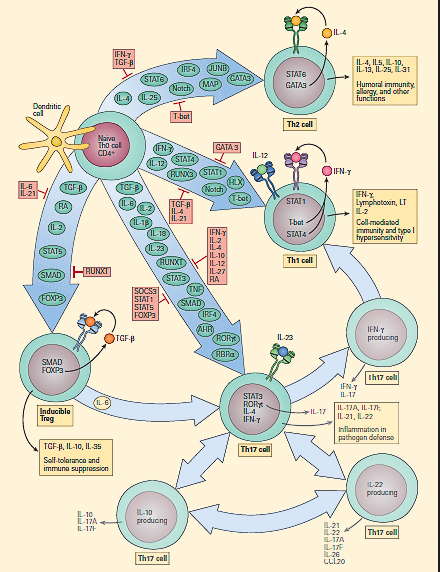
- Treg cells can develop from thymic CD4+ T cell precursors in the presence of TGF-β and IL-2. In the periphery, naïve CD4+ T cells can also be converted to become inducible T reg cells by signaling through STAT5 in the presence of TGF-β, which results in upregulation of the transcription factorforkhead box P3 (FOXP3).
- Treg cells produce high levels of IL-35, CTLA-4, and TGF-β. Retinoic acids, which are abundant in the liver and intestine, increase FOXP3 expression.
- In the presence of commensal bacteria, dendritic cells synthesize retinoic acid from vitamin A, which not only upregulates the FOXP3 transcription factor, but also downregulates the expression of theretinoic acid-related orphan receptors (RORγt and RORα), thereby inhibiting the activation of naïve T cell precursors into Th17 cells.
- Treg cells, therefore, have an important role in peripheral self-tolerance and immune suppression. Th17 cells develop from naïve CD4+ T cells in response to IL-6, IL-1β, IL-23, and TGF-β. IL-1β, IL-6, and IL-23 activate STAT3, which increases the expression of the transcription factors RORγt and RORα, which in turn promote the expression of IL-17A, IL-17F, IL-21, and IL-22.
- IL-23 seems to stabilize and increase the pathogenicity of Th17 cells. Th17 cells are important for host defense against extracellular bacteria and are involved in mediating autoimmune disease. IL-22 tempers the inflammatory effects of IL-17A and IL-17D.
Subset Flexibility and Plasticity
- One of the major hallmark properties of Th differentiation is the amazing capacity of subset flexibility and plasticity rendered by cytokines (FIGURE 4).
- Lineage commitment is defined by the signature cytokines that differentiated cells secrete (i.e., IFN-γ, IL-4, and IL-17, for Th1, Th2, and Th17 cells, respectively).
- However, additional complexity is becoming evident. For example, although Treg cells are sometimes immunosuppressive and produce IL-10, all CD4+ T cell subsets can produce the immunoregulatory cytokine IL-10, which is crucial for dampening immune responses.
- Furthermore, Th17 cells can, at times, also produce IFN-γ, and so the relationship between Th1 cells and Th17 cells is currently being intensively scrutinized.
- Similarly, although Th17 cells produce IL-21 and IL-22, it also seems that under some circumstances, T cells can produce IL-21 or IL-22 without producing IL-17.
- Another limitation adding to the complexity of cytokine secretion by T cell subsets is that proposed models that involve the selective expression of master regulators might not be appropriate. For example, FOXP3 and RORγt can be co-expressed and can interact.
- Moreover, some Treg cells can be induced to become Th17 cells (FIGURE 4). Therefore, Th1 and Th2 CD4+ T cell subsets should be viewed as terminally differentiated lineages, but the Th17 and Treg subsets retain some flexibility and plasticity.
- The extent to which some aspects of T cell subsets are firmly fixed and others remain plastic is the subject of current intense investigation.
Cytokine Receptors and Signaling Pathways
- Cytokines are proteins and key mediators of cellular proliferation, differentiation, and apoptotic cell death and play a central role in both the activation and effector phases of innate and adaptive immune responses.
- All cytokine receptors consist of proteins found at the surface of cells that are composed of three components: (1) an extracellular portion, (2) a transmembrane portion, and (3) an intracytoplasmic portion.
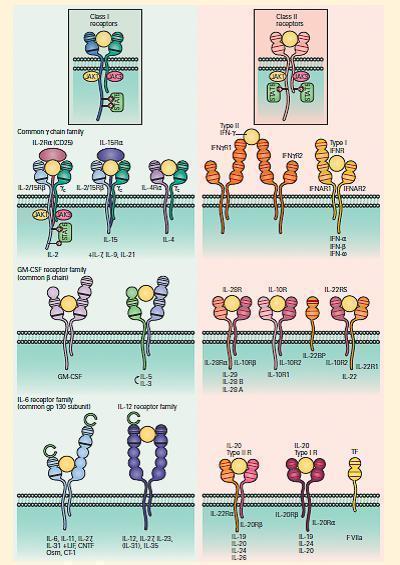
- The cytokine first binds to the extracellular portion of the receptor, following which a signal is generated through a series of conformational changes of the cytokine receptor and a cascading set of biochemical reactions (involving primarily the addition of phosphate groups to substrates, i.e., phosphorylation, through the action of a variety of kinases), during which the signal is transmitted through the cytosolic and nuclear regions of the cell, leading to the ultimate construction of a biologically functional product, e.g., antibody, cytokine.
- The signaling pathways are usually activated by ligand-induced receptor clustering, bringing together the cytoplasmic portions of two or more receptor molecules, and signal transducers and activators of transcription (STATs), described in Table 4.
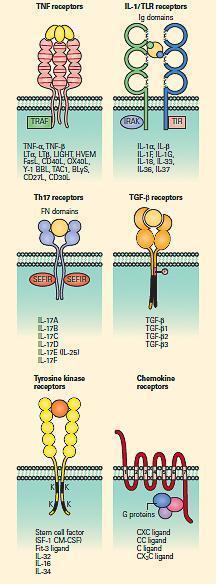
Figure 5B: Schematic representation of the tumor necrosis factor (TNF), IL-1/TLR, Th17, TGF-b serine kinase, tyrosine kinase, and chemokine receptor families. [Reproduced with permission from Bellanti, JA (Ed). Immunology IV: Clinical Applications in Health and Disease. I Care Press, Bethesda, MD, 2012].
- Cytokines and chemokines bind to cell surface receptors that belong to 8 families as shown in FIGURES 5A and 5B and Table 4.
[table id=153 /]
More details on the subfamilies of cytokines, chemokines and their receptors can be found in Bellanti, Immunology IV, Chapter 9.