Activation of B cells
- There are two main immune responses that can drive the activation of resting B-cells, and these are termed as T-cell dependent and independent immune responses.
- Resting B cells become activated by antigen via the BCR and/or by microbiological side products (pathogen associated molecular patterns; PAMPs) via their toll like receptors (TLR4, 7, 9 in mice) and start to proliferate.
- Protein antigens become internalized, digested and presented to T cells as peptides via MHCII.
- Cognate B cell / T cell interaction provides co-stimulation to B cells via CD40, which becomes activated on B cells via CD40 ligand (CD40L) expressed on T cells.
- T cells also provide cytokines to B cells that support their survival (IL-4), differentiation into plasma cells (IL-21) or class switch recombination.
- In mice, Th1 cytokines, such as IFNγ, typical for antiviral responses, elicit IgG2 isotypes, Th2 cytokines, such as IL-4, IL-5 and IL-13, typical for parasite infections, elicits IgG1 and IgE responses.
- With regards to PAMPs, for instance lipopolysaccharide, they elicit differentiation of B cells into short-lived plasma cells secreting low-affinity antibodies. MZ B cells are especially prone to rapidly differentiate into short-lived plasma cells but FO B cells can also differentiate into short- lived plasma cells.
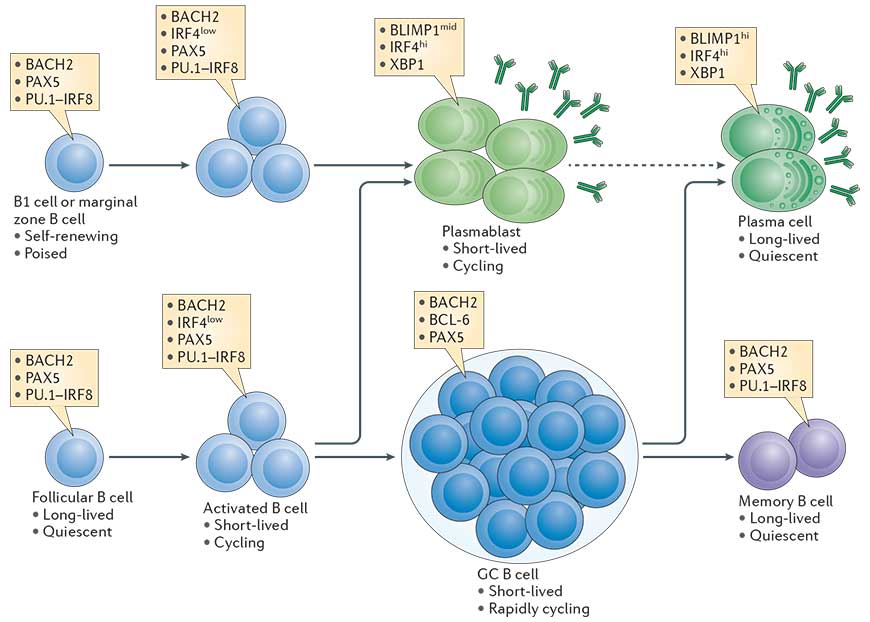
- B cell differentiation into plasma cells is coupled to a certain number of cell divisions that are required to allow expression of transcription factors which terminate the B cell program and initiate the plasma cell program.The transcriptional network that is initiated in B cells in a proliferation dependent manner and fosters plasma cell differentiation is outlined in Fig. 1 (Taken from Nutt et al., Nature Reviews in Immunology, 2015).
- T cell dependent activation of B cells supports the generation of memory B cells and long-lived plasma cells secreting high affinity antibodies. This process requires specific microanatomical structures in secondary lymphoid organs, the germinal centers, where class switch recombination and somatic hypermutation occur.
Precursors of plasma cells
- Plasma cells can be defined as terminally differentiated antibody secreting cells that fall under the B-cell lineage and are non-dividing.
- Resting B2 B cells (these are follicular (FO) and marginal zone (MZ) B cells) express membrane bound IgM, the B cell receptor (BCR) of the IgM type.
- In mice and likely also in humans, another B cell population that is located primarily to the pleural cavities exists, the B1 B cells, a self-renewing population. B1 B cells secrete natural IgM in a T cell independent manner and have a limited repertoire. They differentiate rapidly into short lived plasma cells
- MZ B cells reside in the marginal zone that surrounds the follicles of secondary lymphoid organs and is directly connected to the vasculature. MZ B cells therefore respond to blood born antigens. They react more rapidly to PAMPs and differentiate rapidly into short lived plasma cells
- FO B cells express also membrane bound IgD
- FO B cells reside in the follicles of secondary lymphoid organs and are long lived. They are typically the B cells that interact with cognate T cells.
- Memory B cells express membrane bound Ig of the IgM, IgG or IgA type.
The germinal center (GC) reaction
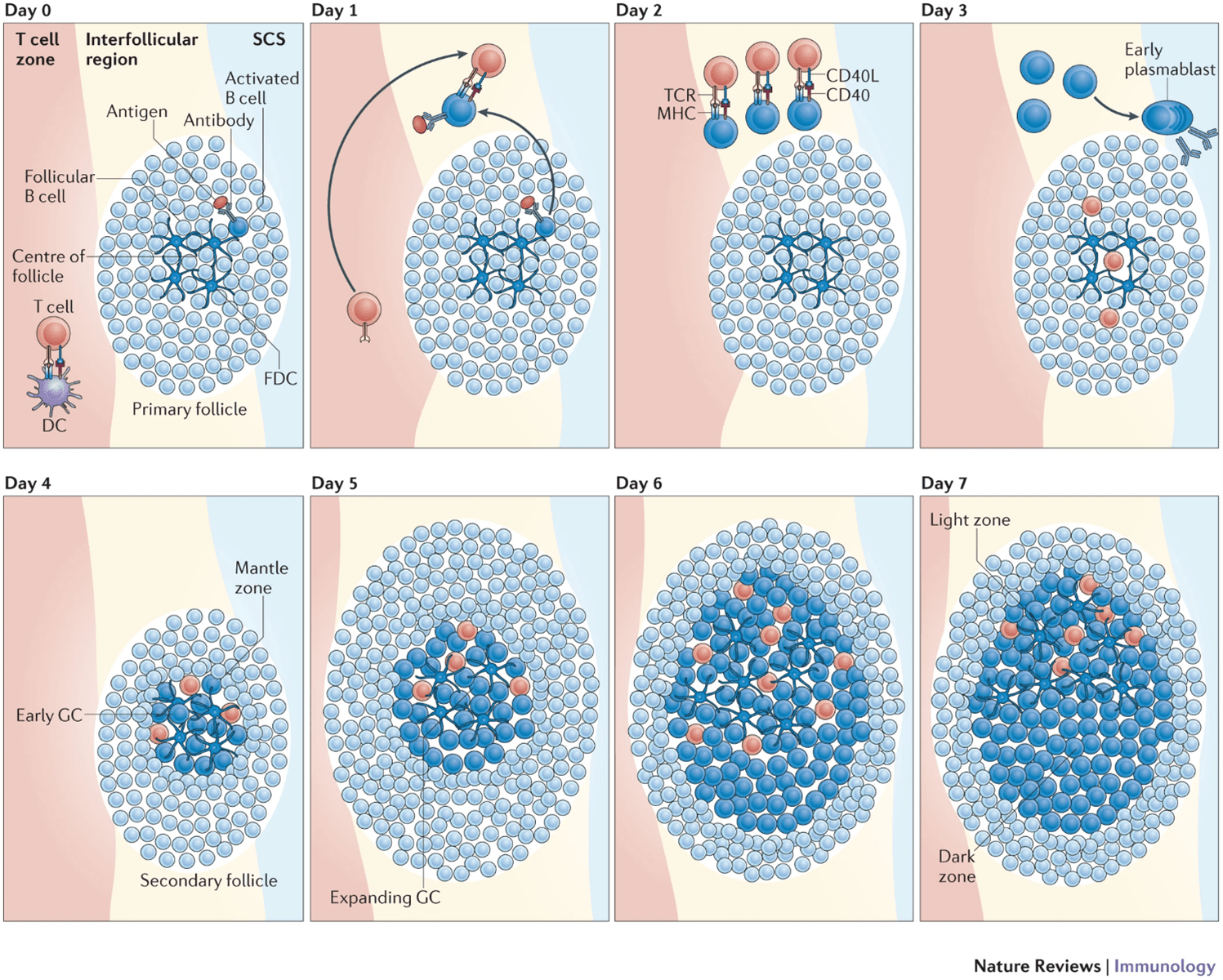
- One of the fundamental pathways needed for the Germinal Center (GC) reaction is the interaction between activated follicular helper CD4 T (Tfh) cells and B cells, highlighted in Figure 2. Tfh cells are essential for the GC reaction as they play a role in increasing the efficiency of antibody responses.
- B-cells encounter antigen in the lymph nodes, thereby triggering their activation. These activated B cells will migrate to the border of the T cell zone where they will encounter T cells. CD40 ligand on T helper cells interacts with CD40 on B cells leading to the secretion of cytokines that will encourage B-cell proliferation and isotype switching.
- GC Tfh cells express IL-21 at high levels. This cytokine is important for the differentiation and functioning of Tfh cells. These cells also produce IL-4, IFN-γ and IL-2 at moderate/variable levels. Bcl-6 is a transcription factor and an important marker of Tfh cells. This transcription factor is important for the development of Tfh and germinal center B cells.
- The survival signals that B-cells receive from T cells allow them to continue proliferating and differentiate into short-lived plasma cells called plasmablasts or form germinal centres (GCs).
- GCs, are specialized regions formed within secondary lymphoid organs and are the sites taken to be responsible for generating mature B-cells, i.e. plasma cells and memory B-cells. GC formation is taken to occur in a T-cell dependant manner due to the same signals that drive B-cell proliferation also being responsible for maintenance of the GC.
- GCs, as shown in Figure 3, are made up of light and dark zones and the sole purpose of GCs is to give rise to antibody-producing plasma cells and memory B-cells with BCRs that have a very high affinity for antigen.
- The GC reaction starts in the dark zone, where B-cells undergo rapid proliferation as well somatic hypermutation (SHM). SHM, initiated by the enzyme activation-induced cytidine deaminase (AID), will lead to random mutations in the variable regions of the BCR. Some of these mutations will result in BCRs that have a larger range of affinities for antigen, thus giving these B-cell clones higher chances of survival when they migrate to the light zone and compete for antigen with other B-cell clones.
- The light zone is taken to be the site where class switch recombination (CSR) starts, however, B-cells will migrate back and forth between both the dark and light zone as they go through multiple rounds of both SHM and CSR.

The plasma cell and antibody secretion
- Early plasma cells still proliferate and are also called plasmablasts or antibody secreting cells (ASC).
- The term “antibody secreting plasma cell” is unambiguous and refers to the terminally differentiated, non-proliferating plasma cell that secretes high amounts of antibody. Plasma cells are much larger than resting or even proliferating B cells.
- In plasma cells and plasmablasts, the exons coding for the membrane domain of Ig molecules of various isotypes have been removed and replaced by exons coding for sequences allowing antibody secretion.
- Plasma cells have a strongly expanded endoplasmic reticulum and express high amounts of molecules involved in antibody folding and secretion.
- Plasma cells can secrete several thousands of antibodies per second.
- Long-lived plasma cells migrate back to the bone marrow where their survival is supported in niches. Stable antibody titers upon vaccination have been observed for 20 years and more for some immunogens.
Quiz
Related Talk
Richard Koup, NIH – Germinal centres and tfh cells