Immunity against Leishmania parasites is mediated by both innate (neutrophils, macrophages, innate like-lymphoid cells and dendritic cells) and adaptive immune mechanisms.1
The host innate immune system represents the first line of defense against invasive pathogens.
Neutrophils: are the first host cells recruited during Leishmania infection within a few hours of inoculation by a sandfly bite.
They have been shown to actively engulf Leishmania promastigotes and produce an array of microbicidal factors against Leishmania such as nitric oxide (NO), neutrophil elastase (NE), platelet activating factor and neutrophil extracellular traps (NETs).2,3
NETs are web-like chromatin structures that play important roles in trapping pathogens; killing pathogens, allowing pathogens to survive and disseminate slowly, and also in non-infectious diseases such as cancer.
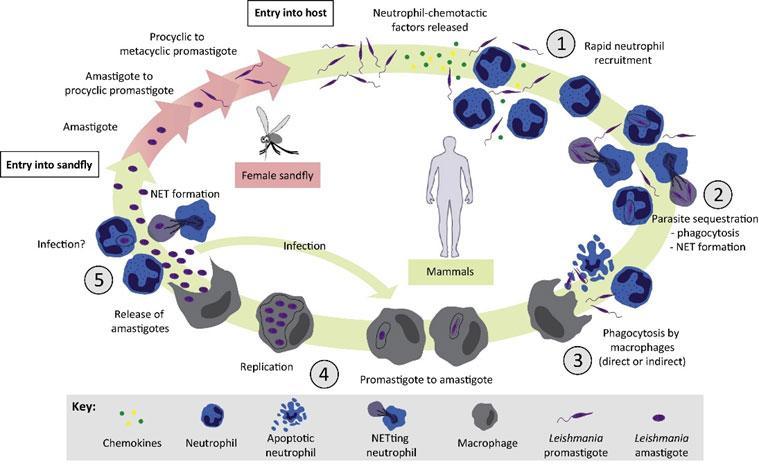
Figure 1: The Role of Neutrophils in the Mammalian Leishmania sp. Life Cycle 4 :(1)Female sandflies transmit infectious metacyclic promastigotes to human or vertebrate hosts through a blood meal. Neutrophils are rapidly and transiently recruited to the site of infection. (2) Parasites are phagocytosed by neutrophils and/or induce the formation of NETs. (3) Leishmania promastigotes directly or indirectly enter macrophages. (4) Promastigotes transform into amastigotes within macrophages and replicate by simple division. (5) Days or weeks after the initial infection, a second wave of neutrophils is recruited to the infection site where they remain during chronic inflammation. Amastigotes are released from bursting macrophages. They will be recaptured by macrophages or by neutrophils. In addition, intralesional parasites can induce NET formation. [Hurrell, B. P., Regli, I. B. & Tacchini-Cottier, F. Different Leishmania Species Drive Distinct Neutrophil Functions. Trends in parasitology 32, 392–401 (2016).]
Innate lymphoid cells (ILCs): have been identified as important sentinel cells at the barrier surfaces that regulate both innate and adaptive immune responses during homeostasis and inflammation. ILCs are divided into:
Group 1 (ILC1): macrophage activating cells.
ILC1s produce protective IFN-γ in response to Leishmania (L.) major (CL) or L. infantum (VL) in an interleukin (IL)-12/IL-18-dependent manner.
NK cells are the primary source of early IFN-γ which favours the Th1 differentiation of CD4+ T cells and restricts early parasite dissemination. NK cells can also mediate direct parasite lysis through their cytotoxic activity and subsequently contribute to cytokine-mediated inducible nitric oxide synthase (iNOS) induction in Leishmania-infected macrophages.
Group2 (ILC2): alternative macrophage activating cells.
ILC2s neither became activated to produce IL-4/IL-13 cytokines nor expand during Leishmania infection, not even in L. major-infected BALB/c mice, which develop systemic disease due to a sustained activity of type 2 T helper (Th2) cells.
Natural killer T (NKT) cells, a specialized subset of T lymphocytes involved in innate immunity against pathogens during the early stages of Leishmania infection.
Group3 (ILC3): produce cytokines typical for the population of Th17 helper lymphocytes and anti-microbial peptides.
The characteristic feature of ILC3s is the expression of the RORγt transcription factor, which is needed for its development and expression of the chemokine receptor CCR6. This group of ILC produces IL-17 (especially IL-17A) and IL-22. ILC3s control epithelial production of antimicrobial peptides whose function is to prevent attachment and translocation of intestinal microorganisms. ILC 3 represent a defence against extracellular parasites and are involved in maintaining intestinal homeostasis5.
IL-17 cytokine family plays an important role in the clearance of intracellular pathogens. In human visceral leishmaniasis (VL) it has been demonstrated that IL-17 and IL-22 are associated with protection against re-exposure to Leishmania6.
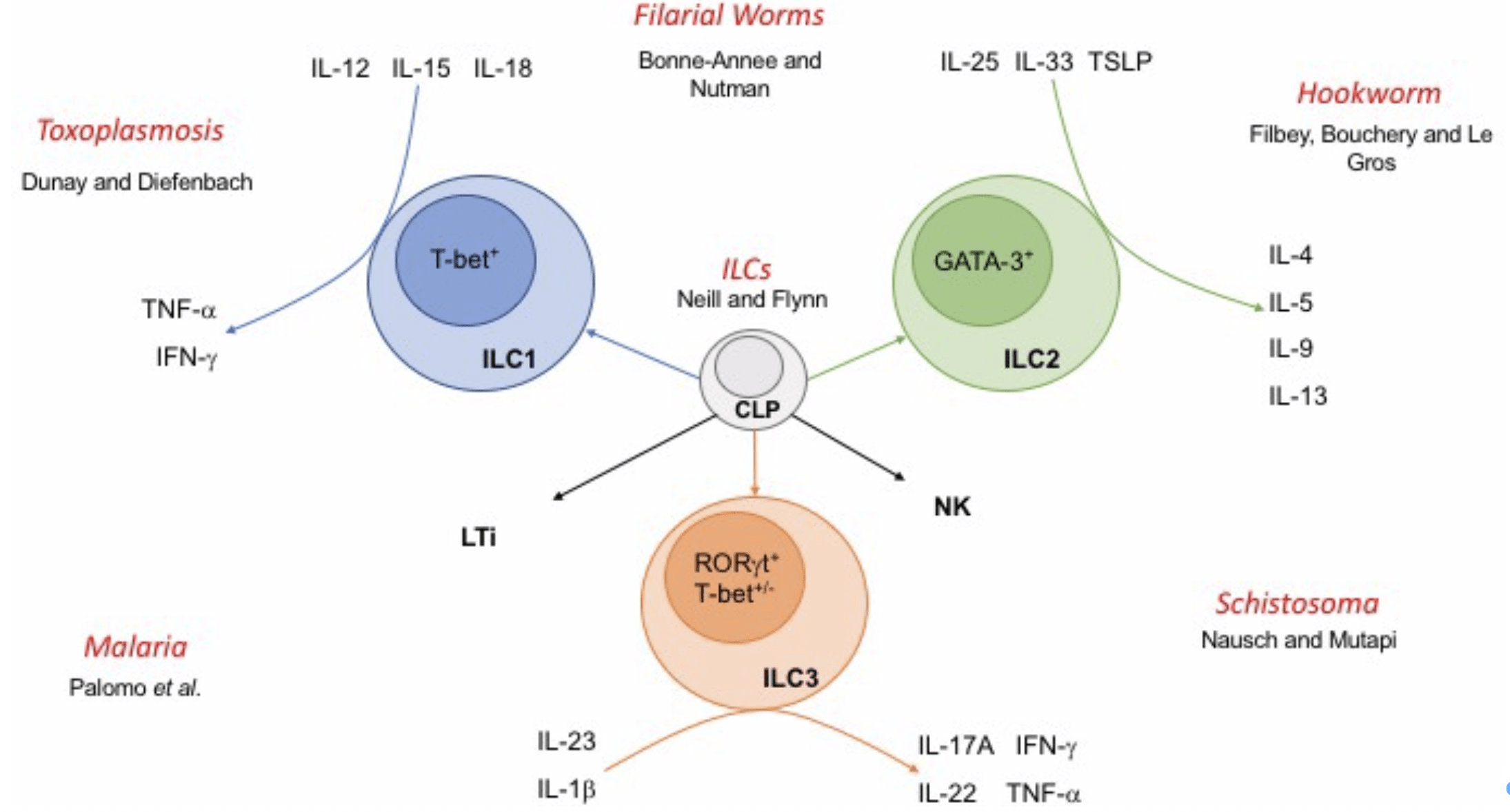
Figure 2: Innate lymphoid cells and their interactions with parasites. Innate lymphoid cells (ILCs) arise from a common lymphoid progenitor (CLP) and are delineated as ILC1, 2 or 3 based upon expression of lineage‐specific transcription factors and production of key cytokines. Natural killer (NK) and lymphoid tissue inducer (LTi) cells also arise from CLP and share some functional characteristics with ILCs. [Neill & Fallon 2018]
A common characteristic of the different innate immune cells is the expression of pattern recognition receptors (PRRs), capable of recognizing pathogen-associated molecular patterns (PAMPs). These PAMPs are recognized via:
cytosolic DNA sensors (CDSs)
NOD-like receptors (NLRs)
RIG-I-like receptors (RLRs)
C-type lectin receptors (CLRs)
Toll-like receptors (TLRs) are expressed intracellularly or at the surface of the cells involved in innate immune responses through recognition of conserved structures on pathogens, such as Leishmania, and play a pivotal role in triggering innate and adaptive immune responses.
TLR2 has a central role in the recognition of lipophosphoglycan (LPG) from L. major and promotes NK cell activation through NFκB activation, culminating in the release of reactive oxygen species (ROS), reactive nitrogens species (RNS) and nitric oxide produced by macrophages. Production of these molecules favour parasite clearance, whereas infection with L. braziliensis promotes the expression of superoxide dismutase (SOD), leading to parasite persistence.
Additionally, TLR2 promotes both activation of DCs and development of the Th1 and Th17 immune response, which together induces neutrophil recruitment to the spleen and liver. In the last instance, neutrophils are activated and produce TNF-α and NO in a TLR2-dependent manner, promoting the restriction of Leishmania parasites.
TLR4 stimulation post-infection promotes TNF-α and NO production, increasing parasite killing, whereas stimulation prior to infection increases IL-10 production, favouring parasite persistence.2
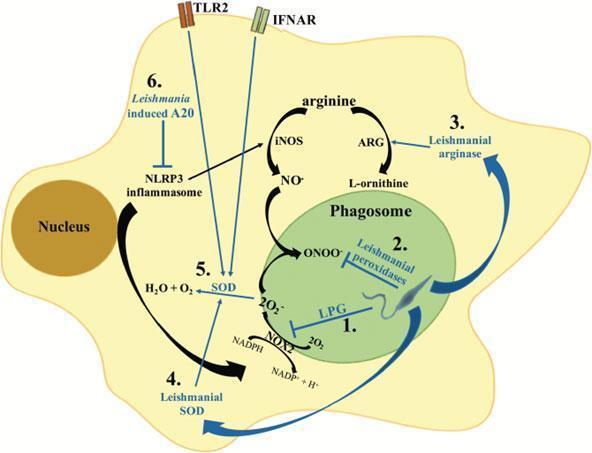
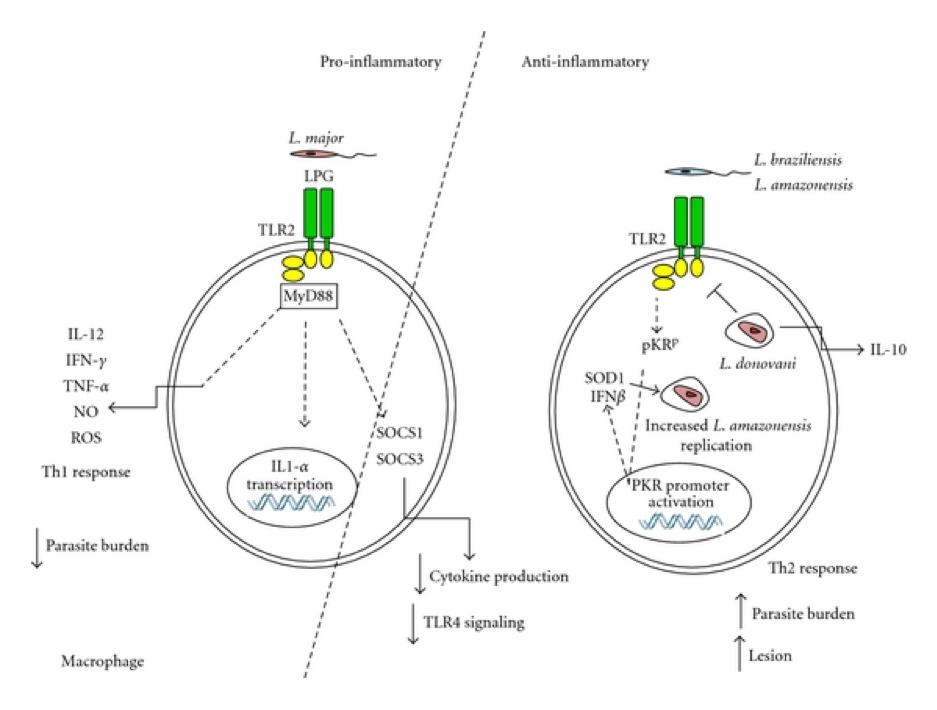
Figure 3: Mechanisms of Leishmania interference with the macrophage oxidative stress response. Leishmania parasites have developed different strategies to interfere with the macrophage oxidative stress response: (1) Early after phagocytosis, the leishmanial LPG coat inhibits the assembly of the NOX2 complex at the phagosome membrane, thus blocking superoxide production (O2–). (2) Leishmania secrete a series of peroxidases which detoxify parasitotoxic peroxynitrite (ONOO–). (3) In addition, Leishmania parasites can secrete arginase. Arginase promotes the conversion of arginine into urea and ornithine required for the parasite metabolism. (4) Secretion of SOD homologues increases the detoxification of parasitotoxic superoxide. (5) Additionally, stimulation of TLR2 or type I interferon receptor (IFNAR) during Leishmania infection induces up-regulation of SOD expression in macrophages. (6) NLRP3 inflammasome activation was shown to promote the production of ROS and RNS; Leishmania inhibits inflammasome activation via the induction of the deubiquitinating protein A20, which inhibits IL-1β maturation. [Rossi, M. & Fasel, N. How to master the host immune system? Leishmania parasites have the solutions! International immunology 30, 103–111 (2018)]Figure 4: Model for the activation of TLR2 by Leishmania sp., leading to a proinflammatory (left), or anti-inflammatory arm (centre and right) [Faria et al. 2012]
Adaptive Immune Response to Leishmania Parasites
CD4+ T cells9
CD4+ Th1 cells are critical for the control of Leishmania infection, as they make IFN-γ that activates macrophages to kill the intracellular parasites3.
Production of IFN-γ results in the upregulation of inducible nitric oxide synthase (iNOS); the latter catabolizes L-arginine into nitric oxide, which is toxic for Leishmania parasites.
CD4+ Th2 cells on the other hand are characterised by the production of cytokines such as IL-4 and IL-13. These cytokines signal via the IL-4 receptor (IL-4R) α chain and both can activate macrophages to upregulate arginase; this enzyme catabolizes L-arginine into polyamines (spermine, spermidine and putrescine) that promote parasite growth. However, it is important to note that both IL-4 and IL-13 dependent and independent factors determine the outcome of experimental L. major infections.
It has been well established that subcutaneous infection of mice with L. major parasites results in different outcomes depending on the strains of mice used: the majority of inbred mice e,g C57BL/6 develop small lesions that heal spontaneously over a few weeks, these mice can control the replication of parasites; in contrast, BALB/c mice develop progressive nonhealing lesions and cannot control parasite replication. This experimental model of Leishmania major infection exemplifies the Th1/Th2 paradigm of resistance/susceptibility to intracellular infection and establishes that the archetypal Th2 cytokines IL-4 and IL-13 counter regulate a protective Th1 response and promote susceptibility to infection.
CD8+ T cells10
Activation of CD8+ T cells during Leishmania infections does not occur via the classical endogenous antigen presentation pathway. It is currently not known how leishmanial peptides are loaded onto MHC class I molecules and how priming of CD8+ T cells occurs in leishmaniasis.
The impact of CD8+ T cells on disease development depends on their effector function: cytotoxic CD8+ T cells can be either beneficial or detrimental to primary L. major infection.
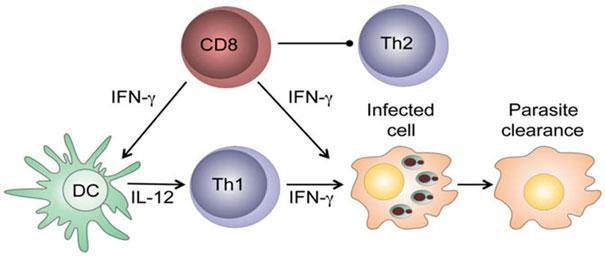
Beneficial effector functions of CD8+ T cells: CD8+ T cells have a protective role in low-dose leishmanial infections: they produce IFN-γ in the draining lymph nodes that can:
activate parasitized macrophages to kill the intracellular parasites.
enhance secretion of IL-12 that promotes the development of CD4+ Th1 cells that can block the development of Th2 cells.
After healing of primary infection, IFN-γ-producing CD8+ T cells contribute to protection against reinfection, are able to transfer delayed type hypersensitivity to recipient mice and are essential in vaccine-induced immunity.
Detrimental effector functions of CD8+ T cells: The cytolytic activity of CD8+ T cells in lesions correlates with disease severity as evidenced by the higher expression of cytolytic markers in the skin lesions of infected susceptible mice as compared to resistant mice.
Figure 5: Protective CD8+ T cells in leishmaniasis. [Novais, F. O. & Scott, P. CD8+ T cells in cutaneous leishmaniasis: the good, the bad, and the ugly, Semin Immunopathol, 2015]
Th17 cells are a subset of activated CD4+ T cells acting as a bridge between adaptive and innate immunity where they promote neutrophil activation, immunity to pathogens and inflammation. In experimental CL, IL-17 has been shown to promote the progression of disease in mice genetically susceptible to L. major infection.
B cells: Mice infected with Leishmania parasites have high levels of parasite-specific IgG. However the role of B cells in experimental CL is controversial, some experiments suggest that B cells can contribute to the control of parasite replication, while other studies show that B cells do not play a role in lesion development or healing. A population of regulatory B cells has been described that was shown to contribute to the development of Th2 cells via the production of IL-1011.
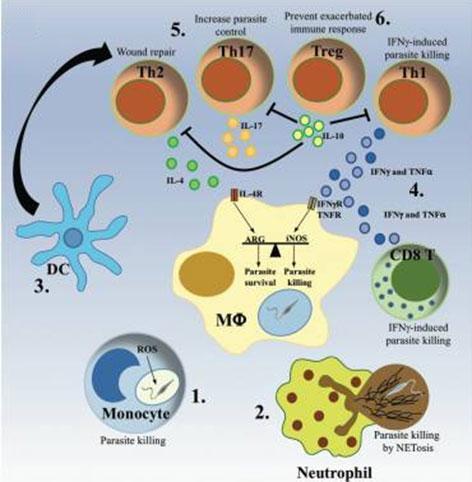
Figure 6: The protective immune response. (1)(2) Monocytes and neutrophils are rapidly recruited to the infection site, where they clear Leishmania parasites via ROS production or NET formation. (3) Dendritic cells (DC) migrate to the lymph node, where they activate and polarize T cells into different Th populations. (4) The development of a predominant Th1 immune response, characterized by production of IFN-γ and TNF-α by CD4+ and CD8+ T cells. (5) Activation of infected macrophages to kill intracellular parasites via production of NO, while a moderate Th2 and Th17 response has been shown to promote the control of the parasite and the resolution of the infection. (6) The subsequent development of IL-10-producing Treg cells ensures the dampening of the immune response, avoiding the development of chronic inflammation. [Rossi, M. & Fasel, N. How to master the host immune system? Leishmania parasites have the solutions! International immunology 30, 103–111]
Kedzierski, L. Leishmaniasis Vaccine. Where are We Today? Journal of global infectious diseases 2, 177–185; 10.4103/0974-777X.62881 (2010).
Rossi, M. & Fasel, N. How to master the host immune system? Leishmania parasites have the solutions! International immunology 30, 103–111; 10.1093/intimm/dxx075 (2018).
Gupta, G., Oghumu, S. & Satoskar, A. R. Mechanisms of immune evasion in leishmaniasis. Advances in applied microbiology 82, 155–184; 10.1016/B978-0-12-407679-2.00005-3 (2013).
Hurrell, B. P., Regli, I. B. & Tacchini-Cottier, F. Different Leishmania Species Drive Distinct Neutrophil Functions. Trends in parasitology 32, 392–401; 10.1016/j.pt.2016.02.003 (2016).
Wenink, Mark H.; Leijten, Emmerik F. A.; Cupedo, Tom; Radstake, Timothy R. D. J. (2017): Review. Innate Lymphoid Cells: Sparking Inflammatory Rheumatic Disease? In: Arthritis & rheumatology (Hoboken, N.J.) 69 (5), S. 885–897. DOI: 10.1002/art.40068.
Pitta, Maira G. R.; Romano, Audrey; Cabantous, Sandrine; Henri, Sandrine; Hammad, Awad; Kouriba, Bouréma et al. (2009): IL-17 and IL-22 are associated with protection against human kala azar caused by Leishmania donovani. In: The Journal of clinical investigation 119 (8), S. 2379–2387. DOI: 10.1172/JCI38813.
Neill, D. R.; Fallon, P. G. (2018): Innate lymphoid cells and parasites. Ancient foes with shared history. In: Parasite immunology 40 (2). DOI: 10.1111/pim.12513.
Faria, Marilia S.; Reis, Flavia C. G.; Lima, Ana Paula C. A. (2012): Toll-like receptors in leishmania infections. Guardians or promoters? In: Journal of parasitology research 2012, S. 930257. DOI: 10.1155/2012/930257.
16 Hurdayal, R. & Brombacher, F. The role of IL-4 and IL-13 in cutaneous Leishmaniasis. Immunology letters 161, 179–183; 10.1016/j.imlet.2013.12.022 (2014).
Novais, F. O. & Scott, P. CD8+ T cells in cutaneous leishmaniasis. The good, the bad, and the ugly. Seminars in immunopathology 37, 251–259; 10.1007/s00281-015-0475-7 (2015).
Ronet, C. et al. Regulatory B cells shape the development of Th2 immune responses in BALB/c mice infected with Leishmania major through IL-10 production. J Immunol 184, 886-894, doi:10.4049/jimmunol.0901114 (2010).