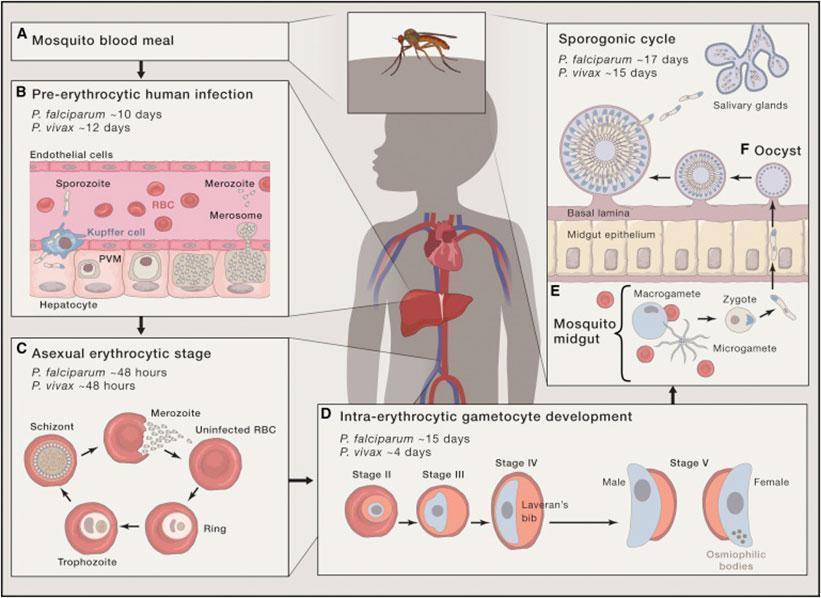
Regarding vaccine development there exist several approaches, as immune responses could potentially target any stage of the parasite to which humans are exposed (Fig. 1)
Figure 1: Life cycle of P. falciparum. (A) During a mosquito blood meal Plasmodium sporozoites are injected into the dermis.(B) Through the vasculature the sporozoites are transported to the liver where they enter hepatocytes and develop within 7-10 days (P. falciparum) to schizonts which release tens of thousands merozoites into the vasculature.(C) The merozoites infest erythrocytes and perform a cycle of asexual schizogony by developing into trophozoites and subsequently schizonts which release merozoites.(D) A small part of merozoites develop into the sexual stages of the parasite, the male (microgamete) and female (macrogamete) gametocytes.(E) When a mosquito takes a blood meal containing both micro- and macrogametes, they emerge in the midgut of the mosquito and mate, thereby forming a zygote which transforms into an ookinete and finally into an oocyst (F) which releases, when mature, sporozoites which migrate into salivary glands where they can be transferred to a human host during next blood meal.[adapted from Cowman, A.F., et al., Malaria: Biology and Disease. Cell, 2016. 167(3): p. 610-624.]Pre-erythrocytic vaccines (PEs)
The clinical vaccine development, however, has focused on the pre-erythrocytic stage which comprises sporozoites and liver stages of P.falciparum
Full protection would be achieved through stopping establishment of blood-stage infection and thereby preventing a clinical malaria infestation
For this both, T-cells and antibodies are needed to eradicate infected hepatocytes, target parasites and prevent sporozoites to enter the liver [4]
CD8+ and CD4+ T cells are required, as well as memory T cells for long term protection
The only licensed malaria vaccine so far is RTS,S [5], a PE that targets circumsporozoite (CS) protein, which is the major coat protein of sporozoites
It partly consists of the C-terminal end of P.falciparum CS which is fused with a hepatitis B surface antigen virus like particle (VLP)
CS-specific antibodies and CD4+ T cells are responsible for protection whereas CD8+ T cells play a minor part [6]
Unfortunately, the efficacy was poor in clinical trials, as only 28% of children aged 5-17 months were protected after three vaccine doses (at study end, after 48 months) [7]. After 18 months, efficacy was higher with 45% for 5-17 months old children.
Moreover, the protective antibodies wane over time, leading to a short duration of protection
Another approach is the combining of RTS,S with other malaria vaccine regimens to achieve improving of efficacy by eliciting more CD8+ T cells : The Chimp Adenovirus (ChAd)-MVA Prime Boost strategy in which probands are primed with chimpanzee adenovirus 63 (ChAd63) and boosted 8 weeks later with modified vaccinia virus Ankara (MVA) [8]
Highly immunogenic, inducing CD4+ and CD8+ T cells as well as anti-CS antibodies
But, sterile protection only in 1 out of 15 individuals [9]
Similar results using the antigen TRAP (thrombospondin related adhesion protein) instead of CS; it is safe and immunogenic but induced sterile protection in only 2 out of 15 individuals [9]
Use of radiation-attenuated, cryopreserved whole P.falciparum sporozoites (PfSPZ) as a vaccine, is another promising trial
After infesting hepatocytes, irradiated sporozoites do not initiate progress to blood stage development, if sterile protection is achieved [10]
PfSPZ vaccine induces antibodies and effector and memory T cells and is well tolerated [11, 12]
Sterile protection up to 100% (for > 1 year) was achieved in clinical trials after homologous 3-5 times CHMI (controlled human malaria infection) [13, 14]
Big challenges are the complexity of Plasmodium with its highly polymorphic antigens, resulting in problems with achieving heterologous protection (cross-strain or strain-transcendent protection) after vaccination, as well as the need of mass production of sporozoites and the obligation to maintain a liquid nitrogen cold chain for the parasites in remote areas
Pregnancy-associated malaria
P.falciparum parasites accumulating in the placental tissue during pregnancy cause placental malaria (PM)
PM is mostly asymptomatic but is associated with low birth weight or stillbirth and maternal anemia
Infection leads to almost a million low birth weight deliveries in Africa each year [15]
P. falciparum infected erythrocytes bind to chondroitin sulfate A (CSA), a glycosaminoglycan composed of disaccharides of N-acetylgalactosamine and D-glucuronic acid, fused to placental proteoglycans
Main proteoglycan responsible for parasite adhesion in the placenta is Syndecan-1 [16]
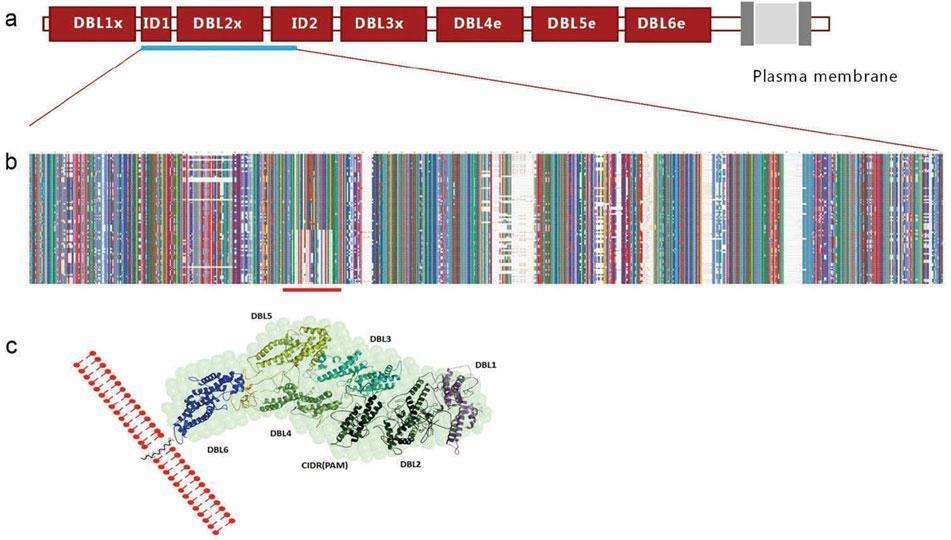
A study from 2003 identified the protein VAR2CSA, which is expressed on the surface of infected erythrocytes, to be associated with the binding to CSA [17] (Fig. 2)
Figure 2: Schematic structure of VAR2CSA with the CSA binding region underlined in blue [adapted from Pehrson, C., et al., Pre-clinical and clinical development of the first placental malaria vaccine. Expert Rev Vaccines, 2017. 16(6): p. 613-624.]
In 2004 it was shown that the infants born to women with naturally acquired high amounts of protective antibodies – specific against VAR2CSA – had a higher birth weight compared to those babies born to women with low antibody levels [18]
The VAR2CSA specific antibodies are associated with preventing parasite-infected erythrocytes from binding to CSA in the placenta
Therefore, a VAR2CSA-based vaccine would be a promising trial, as also strategies to prevent malaria in pregnancy are rather not successful: the use of insecticide-treated bed-nets (ITNs) during pregnancy is lower than 40% in sub-Saharan Africa and the administration or rather taking of intermittent preventive treatment in pregnancy (IPTp) with sulfadoxine-pyrimethamine (SP) is estimated to be only 17% in sub-Saharan Africa [19]
So far, a VAR2CSA-based vaccine has reached clinical phase I trials in Europe and Africa [20]
The vaccine candidate contains the N-terminally located CSA binding domains of VAR2CSA and should be administered before pregnancy because the antibodies should be present in maternal plasma before maternal circulation is established in the placenta
Challenges in the vaccine development are the sufficiently high and long-lasting responses, which are required because the vaccine will be administered before pregnancy, the non-protectiveness against non-falciparum malaria and the polymorphism of the antigen
Wilson, K.L., et al., Malaria vaccines in the eradication era: current status and future perspectives. Expert Rev Vaccines, 2019. 18(2): p. 133-151.
Rts, S.C.T.P., Efficacy and safety of RTS,S/AS01 malaria vaccine with or without a booster dose in infants and children in Africa: final results of a phase 3, individually randomised, controlled trial. Lancet, 2015. 386(9988): p. 31-45.
Cockburn, I.A. and R.A. Seder, Malaria prevention: from immunological concepts to effective vaccines and protective antibodies. Nat Immunol, 2018. 19(11): p. 1199-1211.
Greenwood, B. and O.K. Doumbo, Implementation of the malaria candidate vaccine RTS,S/AS01. Lancet, 2016. 387(10016): p. 318-319.
de Barra, E., et al., A phase Ia study to assess the safety and immunogenicity of new malaria vaccine candidates ChAd63 CS administered alone and with MVA CS. PLoS One, 2014. 9(12): p. e115161.
Hodgson, S.H., et al., Evaluation of the efficacy of ChAd63-MVA vectored vaccines expressing circumsporozoite protein and ME-TRAP against controlled human malaria infection in malaria-naive individuals. J Infect Dis, 2015. 211(7): p. 1076-86.
Doolan, D.L. and N. Martinez-Alier, Immune response to pre-erythrocytic stages of malaria parasites. Curr Mol Med, 2006. 6(2): p. 169-85.
Epstein, J.E., et al., Live attenuated malaria vaccine designed to protect through hepatic CD8(+) T cell immunity. Science, 2011. 334(6055): p. 475-80.
Luke, T.C. and S.L. Hoffman, Rationale and plans for developing a non-replicating, metabolically active, radiation-attenuated Plasmodium falciparum sporozoite vaccine. J Exp Biol, 2003. 206(Pt 21): p. 3803-8.
Epstein, J.E., et al., Protection against Plasmodium falciparum malaria by PfSPZ Vaccine. JCI Insight, 2017. 2(1): p. e89154.
Ishizuka, A.S., et al., Protection against malaria at 1 year and immune correlates following PfSPZ vaccination. Nat Med, 2016. 22(6): p. 614-23.
Walker, P.G., et al., Estimated risk of placental infection and low birthweight attributable to Plasmodium falciparum malaria in Africa in 2010: a modelling study. Lancet Glob Health, 2014. 2(8): p. e460-7.
Ayres Pereira, M., et al., Placental Sequestration of Plasmodium falciparum Malaria Parasites Is Mediated by the Interaction Between VAR2CSA and Chondroitin Sulfate A on Syndecan-1. PLoS Pathog, 2016. 12(8): p. e1005831.
Salanti, A., et al., Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol Microbiol, 2003. 49(1): p. 179-91.
Salanti, A., et al., Evidence for the involvement of VAR2CSA in pregnancy-associated malaria. J Exp Med, 2004. 200(9): p. 1197-203.
van Eijk, A.M., et al., Coverage of intermittent preventive treatment and insecticide-treated nets for the control of malaria during pregnancy in sub-Saharan Africa: a synthesis and meta-analysis of national survey data, 2009-11. Lancet Infect Dis, 2013. 13(12): p. 1029-42.
Institut National de la Santé Et de la Recherche Médicale, F., Trial to Evaluate the Safety and Immunogenicity of a Placental Malaria Vaccine Candidate (PRIMVAC) in healthy adults (PRIMALVAC). clinicalTrials.gov [Internet]. Bethesda (MD): National Library of Medicine (US), 2000.